vision-research.eu - The Gateway to European Vision Research
![]()

![]()
You are here: vision-research.eu » Vision Research » The Young Researchers View » The Young Researcher's View: Jenna Nagy (Q4-2021)
The Research of Jenna Nagy
 |
The retina is the brain’s window to a diverse and dynamic visual world with luminance intensities that can vary more than one billion-fold between day and night. Even within a single natural scene, the intensity of luminance can vary by a factor of more than ten thousand. Such dynamic changes in environmental light stimuli are compounded as animals sample their visual environments through exploration, saccades, and eye fixation. Visually guided behavior depends on the retina’s ability to encode these vast stimuli with limited neural machinery. Retinal circuits meet these demands by matching their input-output relationship to the light intensity statistics of natural scenes through the process of adaptation. As an early career retinal neurobiologist, I am interested in how cellular and circuit mechanisms shape light adaptation to a wide range of visual inputs in diverse retinal circuits.
A common mechanism that is particularly apt to mediate adaptation in sensory circuits through adjustment of gain and kinetics is synaptic inhibition. Inhibition regulates neural circuit function by dynamically modulating neuronal excitability, which tunes neurons to specific stimuli and controls important aspects of their responses. Inhibition is mediated by neurotransmitters - GABA or Glycine. Presynaptic inhibition is a widespread, but elusive process in the central nervous system that suppresses transmitter release at the axon terminals of presynaptic neurons, thus controlling the excitatory input received by postsynaptic neurons. A crucial site in the retina where presynaptic inhibition plays a key role in shaping visual signals is at the axon terminals of second-order bipolar cells (BCs) before the BC signal is transmitted to ganglion cells (GCs) which convey retinal output to the brain (Fig. 1A).
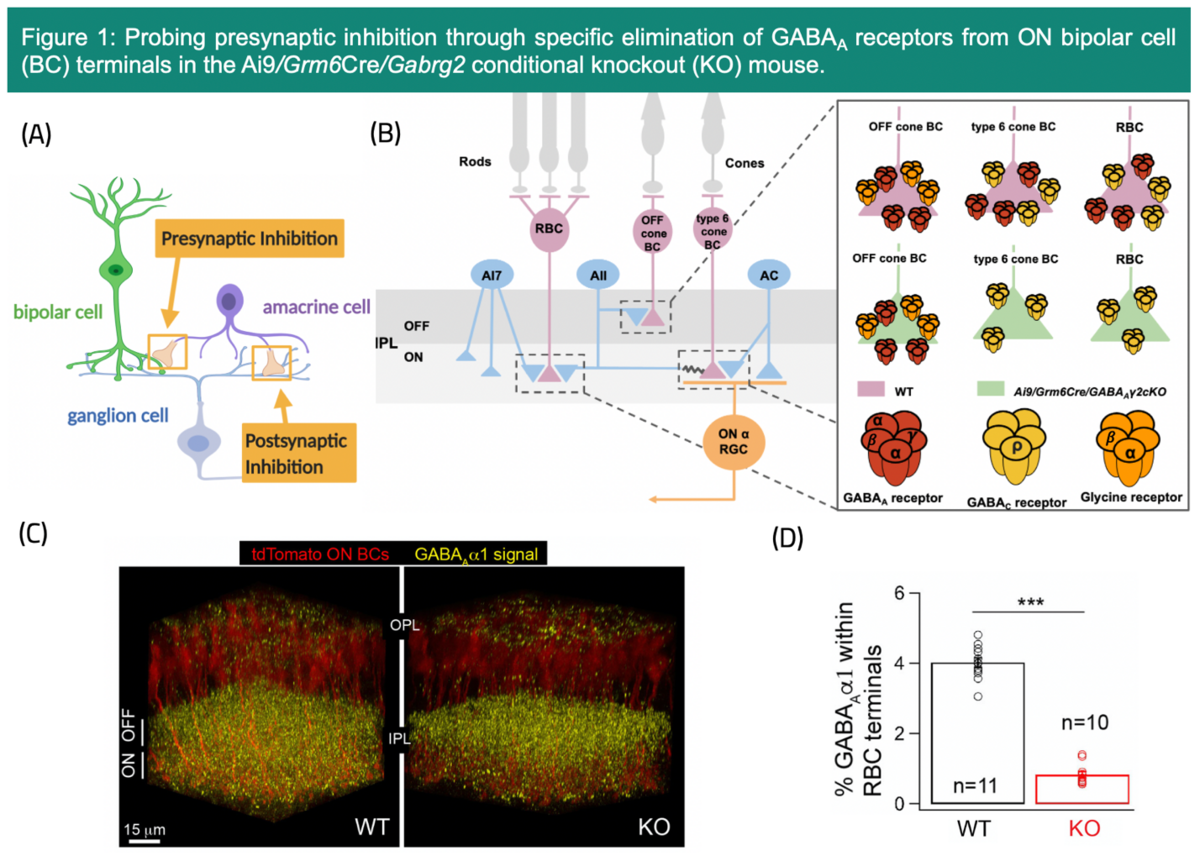 |
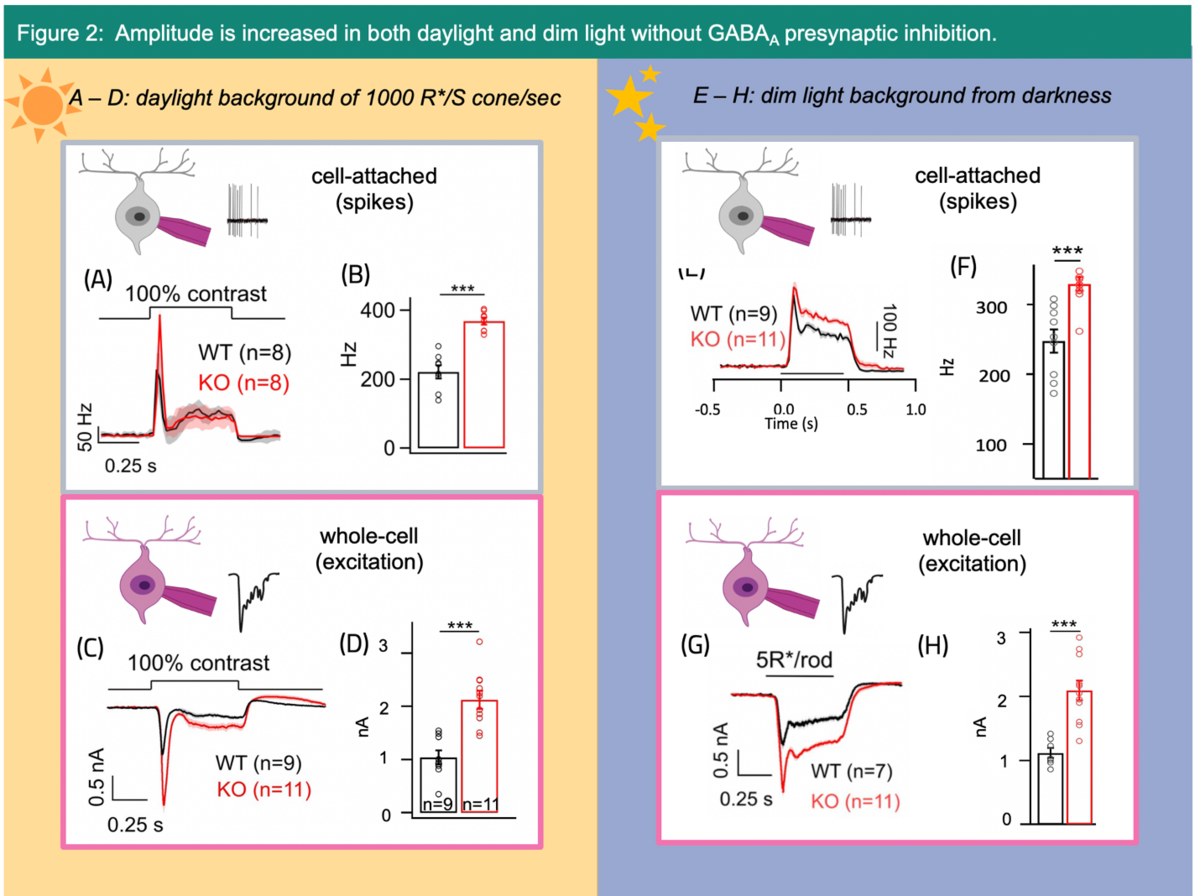
Despite being a central mechanism, technical limitations have made the function of presynaptic inhibition poorly understood. As a PhD student in the lab of Dr. Raunak Sinha at the University of Wisconsin-Madison, USA, I have had a unique opportunity to study the impact of presynaptic inhibition with unprecedented precision by utilizing a novel transgenic approach1, single cell electrophysiology and optical imaging techniques. Previous transgenic mouse approaches targeted the entire retinal circuitry, precluding insights into the role of presynaptic inhibition in specific circuits because these mice globally lacked GABAA receptors, which are densely expressed throughout the inner synaptic layer (Fig. 1C).2, 3, 4 To increase resolution, our lab utilized a unique transgenic mouse line that selectively eliminates GABAA type inhibitory receptors from the axon terminals of two specific types of BCs (Rod BC and type 6 BC) (Fig. 1B)1, 4. These cells are part of the well-characterized ONɑ GC circuit that routes both dim and daylight signals to the brain. In our recent publication, we found that GABAA presynaptic inhibition enhances the sensitivity of the light-evoked responses of ONɑ GCs in both dim and daylight circuits (Fig. 2).4
 |
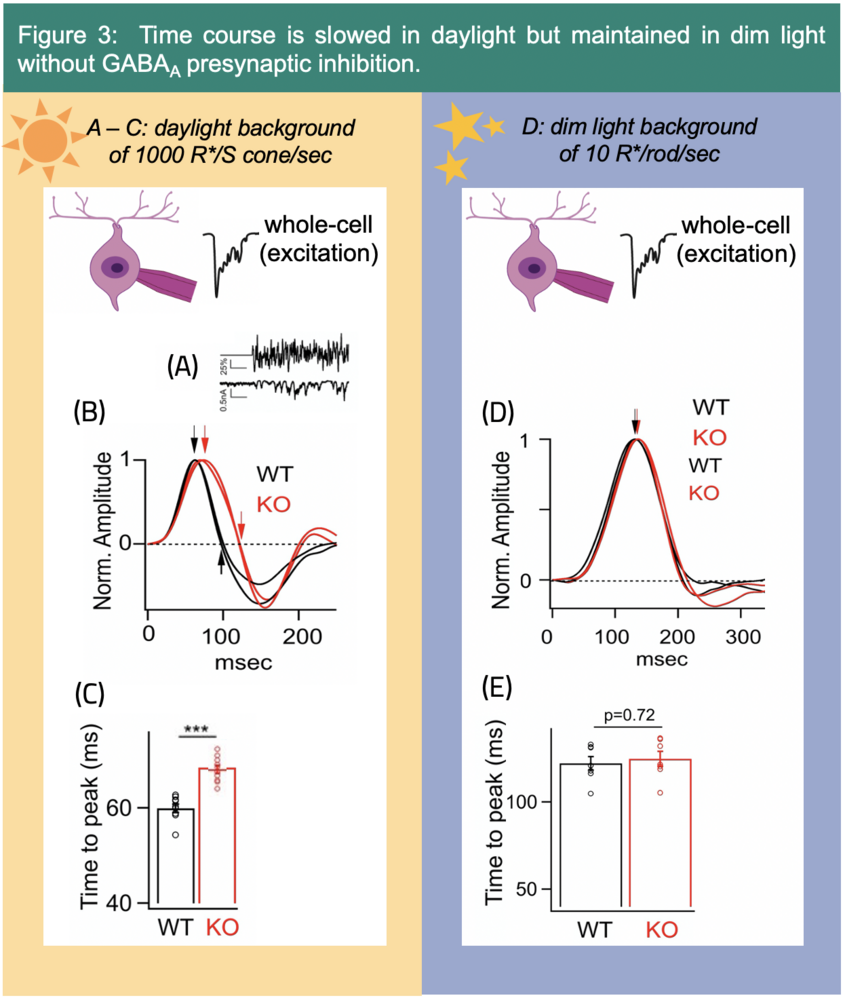
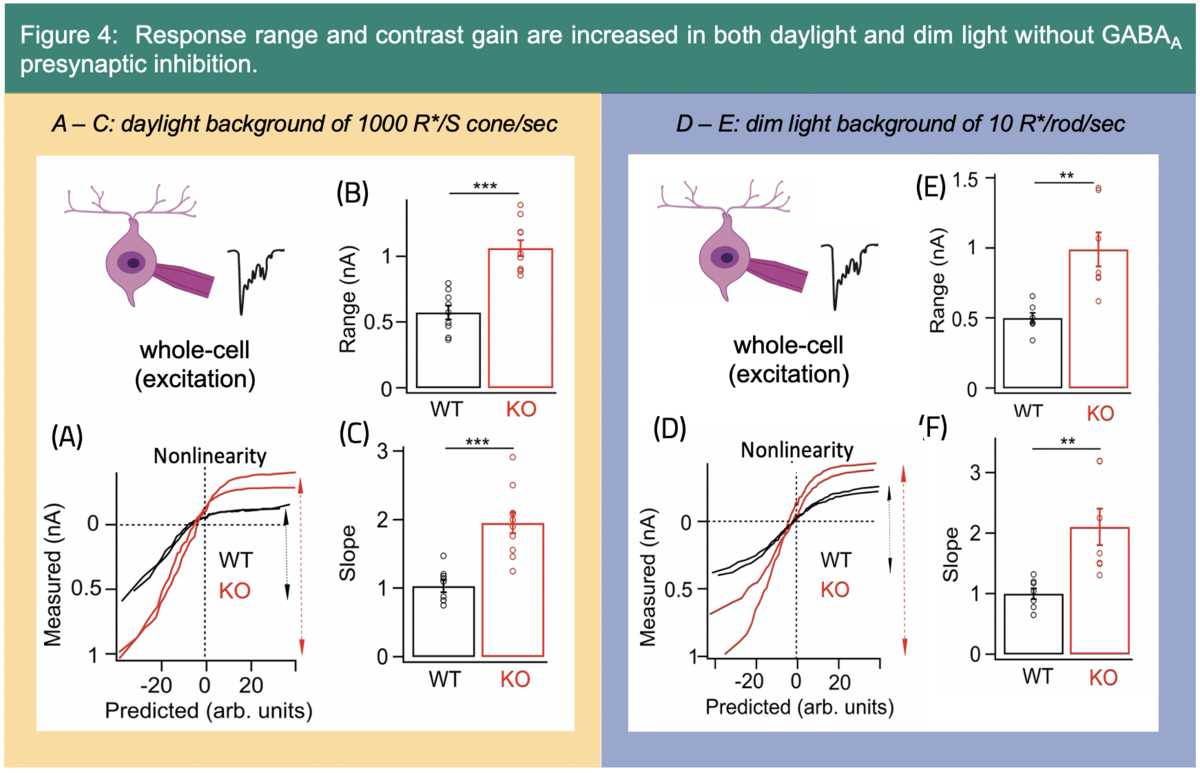
Additionally, GABAA mediated presynaptic inhibition shapes the response kinetics (Fig. 3) and contrast gain of ONɑ excitatory input (Fig. 4).4 Interestingly, GABAA presynaptic inhibition speeds up the time course of signaling only in the daylight circuit.
 |
 |
Our findings highlight the role of presynaptic inhibition in temporal filtering and gain control in a key neural circuit in mouse retina. Our results suggest a potential role for GABAA presynaptic inhibition in ONɑ GC circuit adaptation to major statistics of light input: mean luminance and contrast. Future studies will probe a much wider range of background luminances and contrasts to represent more closely those present in natural scenes. When combined with behavioral approaches, this work will provide a more expansive investigation of the precise roles of presynaptic inhibition in adaptation and visually guided behavior.
References
- Hoon M, Sinha R, Okawa H, Suzuki SC, Hirano AA, Brecha N, Rieke F, Wong RO. Neurotransmission plays contrasting roles in the maturation of inhibitory synapses on axons and dendrites of retinal bipolar cells. Proc Natl Acad Sci U S A. 2015 Oct 13;112(41):12840-5. doi: 10.1073/pnas.1510483112. Epub 2015 Sep 29. PMID: 26420868; PMCID: PMC4611619.
- Eggers, E.D., and Lukasiewicz, P.D. (2011). Multiple pathways of inhibition shape bipolar cell responses in the retina. Vis Neurosci 28, 95-108.
- Oesch, N.W., and Diamond, J.S. (2019). Synaptic inhibition tunes contrast computation in the retina. VisNeurosci 36, E006.
- Nagy, J., Ebbinghaus B, Hoon M, Sinha R. GABAA presynaptic inhibition regulates the gain and kinetics of retinal output neurons. Elife. 2021 Apr 27;10:e60994. doi: 10.7554/eLife.60994. PMID: 33904401; PMCID: PMC8110304.

Jenna Nagy
Department of Neuroscience, University of Wisconsin, Madison (UWM), United States
McPherson Eye Research Institute, UWM
Cellular and Molecular Pathology Training Program, UWM
E-mail: jnagy2[at]wisc.edu