vision-research.eu - The Gateway to European Vision Research
![]()

![]()
You are here: vision-research.eu » Vision Research » Visionary of the Quarter » Meg Veruki and Espen Hartveit (Q03-2021)
The research work of Professors Meg Veruki and Espen Hartveit
 |
 |
Revealing the relationship between structure and function
Our lab is interested in the cellular and molecular basis of synaptic transmission and synaptic integration in the central nervous system, using the retina as an ideal model system. Whereas the human brain is enormously complex, with estimates of 100 billion neurons, it our hope that the retina with only 100 million neurons might be slightly easier to understand. We study the synaptic and extrasynaptic connections between different types of neurons, each with unique structural and functional properties, that together form the neural microcircuits that process visual signals, e.g. the rod pathway microcircuit illustrated in Figure 1. The primary techniques we use are targeted patch-clamp recording and live-cell injections of visually-identified neurons in acute retinal slices, immunolabeling, confocal microscopy, 2-photon structural and functional imaging, and computational modeling.
, AII amacrine (yellow), rod bipolar cell (RBC; medium blue), OFF-cone bipolar cell (OFF-CBC; light blue), and ON-cone bipolar cell (ON-CBC; dark blue). A Müller glial cell (light red) is also included. Photoreceptors are omitted for simplicity. Adapted from Beltrán-Matas et al., 2021.") |
The AII amacrine cell
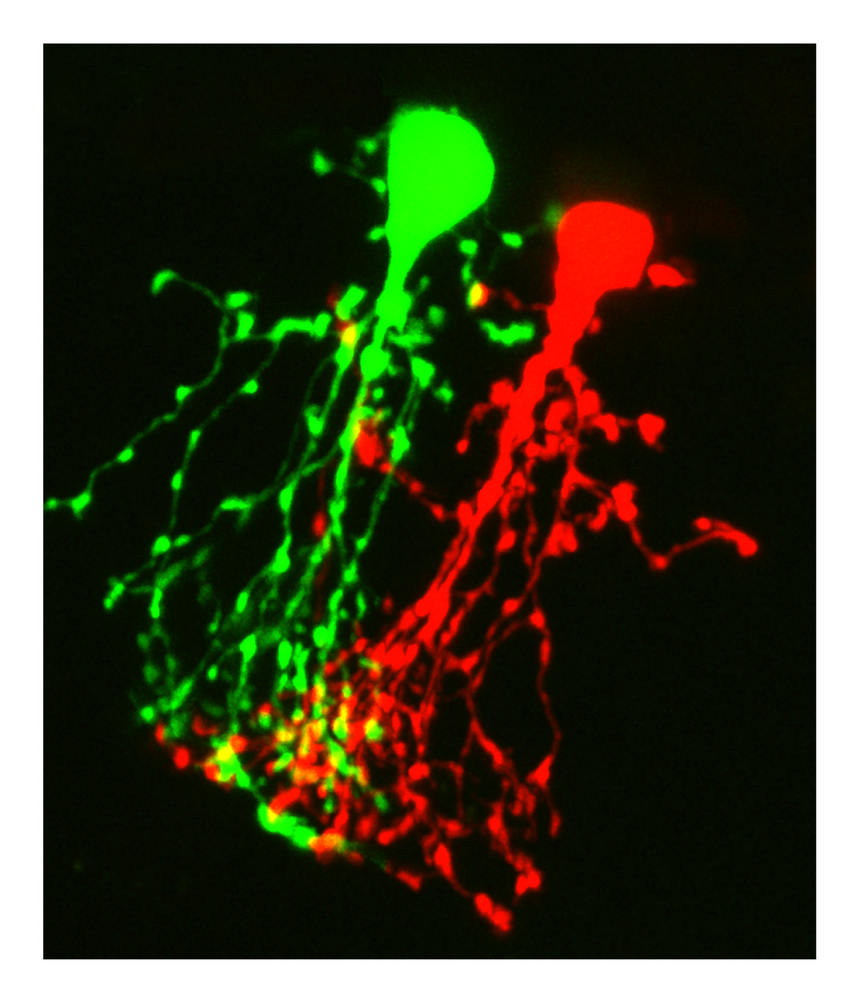 |
Much of our work has been focused on the AII amacrine cell, an intriguing, multifunctional interneuron that seems to poke its processes into all aspects of vision. We have characterized both glutamatergic and glycinergic inputs to AIIs, as well as the electrical synapses that couple these neurons to each other and to ON-cone bipolar cells. Our first paper together was a short report about NMDA receptors in the AII amacrine cell (Veruki and Hartveit, 1997). The paper was neglected by us and others for several years, but we recently turned our attention back to these receptors to discover that they are purely extrasynaptic. We have characterized the molecular composition and location of these receptors on the AII (Zhou et al., 2016; Veruki et al., 2019), and most recently we demonstrated that they are activated by spillover of synaptically released glutamate (Beltran-Matas et al., 2021). These receptors are likely to play a major role, along with dopamine, in modulating the gap junction coupling of these neurons. Although gap junctions between adjacent AIIs and between AIIs and ON-cone bipolar cells had been identified with electron microscopy and studied with intercellular diffusion of tracers, we were the first to demonstrate, using dual patch-clamp recording, that the gap junctions formed functional electrical synapses (Veruki and Hartveit 2002a, b) that transfer visual signals and reduce noise. We went on to characterize the dynamic range and consequences of variations in the strength of coupling, and verified that the gap junction blocker, meclofenamic acid could be used as a pharmacological tool to block these connections (Veruki et al. 2008; Veruki and Hartveit, 2009). Along the way, we found that the AII (as well as the A17 amacrine) are compromised during the early stages of diabetic retinopathy (Castilho et al., 2015a, b). Current projects in our laboratory are to investigate how glutamate and dopamine might regulate the strength of gap junction coupling between AII amacrine cells under different light conditions.
Morphologically realistic compartmental models to elucidate structure function relationships of signal integration
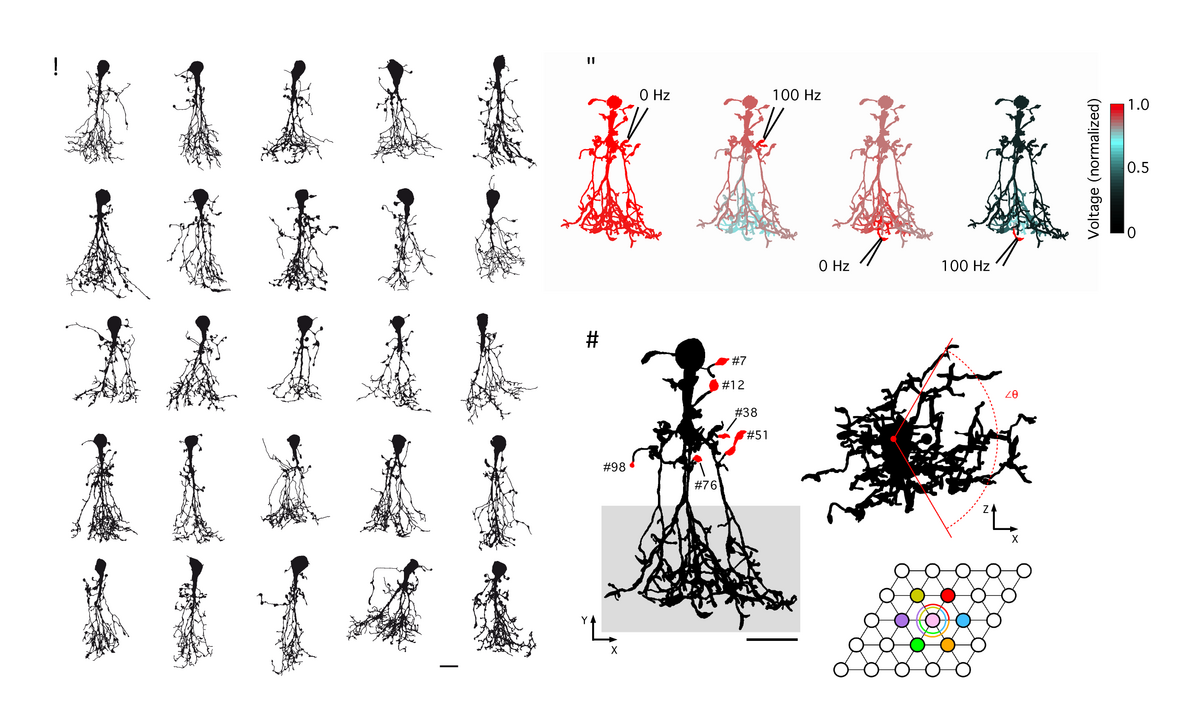
The properties that shape signal integration of a given neuron are determined by both its morphology and physiological characteristics. Recently, we have combined 2-photon microscopy with patch-clamp electrophysiology to develop morphologically realistic compartmental models of AII amacrines (Zandt et al., 2017, 2018). Our detailed quantitative morphological analysis of digitally reconstructed AIIs demonstrates an unexpectedly complex branching pattern, not appreciated from earlier descriptions using conventional wide-field light or electron microscopy, and establishes a database of morphological properties as a reference for future studies of development, degeneration and disease processes. From these reconstructions we developed compartmental models that allow a thorough investigation of the relationship between the structure and function of this complex and fascinating cell. A major result of our initial study is that AII amacrines, despite their relatively small size, cannot be characterized as electrotonically compact. Their morphology and passive properties contribute significantly to signal integration and processing. We found significant frequency-dependent attenuation which was most pronounced for signals generated at the arboreal dendrites and propagating towards the soma and lobular dendrites. We have applied these models to explore the validity of capacitance measurements of exocytosis from the lobular dendrites, including under varying conditions of electrical coupling (Hartveit et al., 2019). In the future, these single cell models will serve as the building blocks for anatomically realistic retinal network models.
 |
Axon initial segment (AIS) in an axon-less amacrine
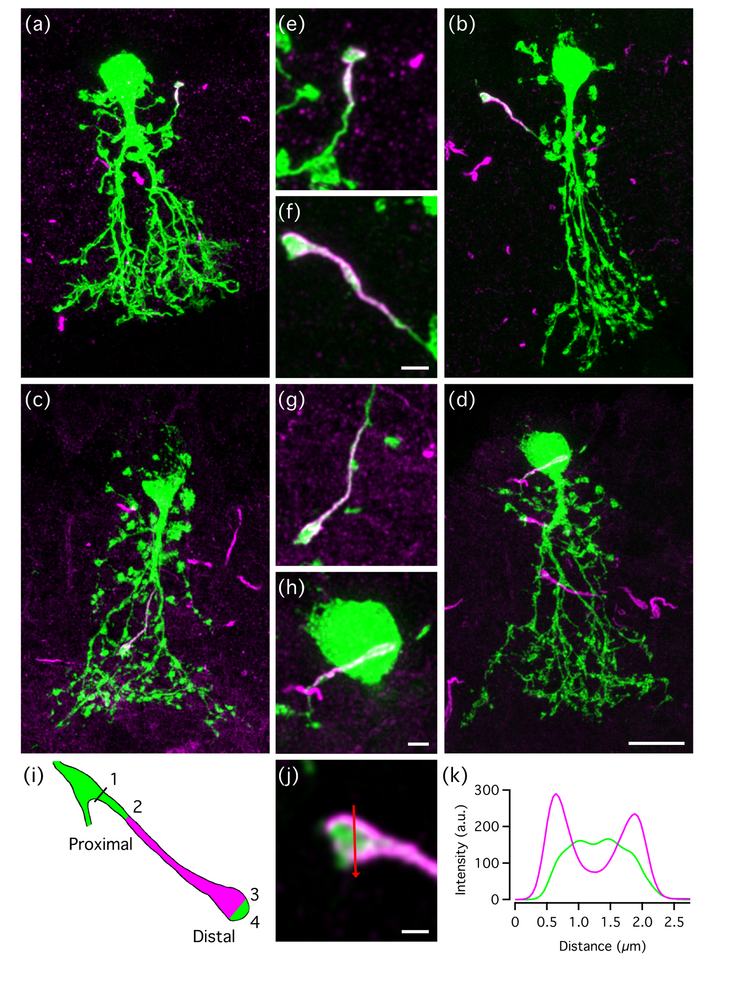
Recent investigations from several laboratories have revealed the presence of an axon initial segment (AIS)-like process on AII amacrine cells. As a first step to investigate the role of spiking in the AII, we systematically characterized the morphology, laminar localization, spatial orientation and distribution of this process (Liu et al. 2021). Using a combination of live-cell injections in retinal slices and immunohistochemistry and confocal imaging of wholemount retina, we investigated the properties of this process as a function of eccentricity. In general, the level of stratification of processes in the inner plexiform layer of the retina defines the functional connectivity of a given neuron. However, we found that across the retina, the AIS-like process on AII amacrines terminated in an essentially random fashion in the inner plexiform layer and could even ascend into the inner nuclear layer. These properties suggest that the AIS-like process might act as an antenna to sense levels of a volume neurotransmitter by means of extrasynaptic receptors.
 |
Not only AII amacrines
In addition to our work on the AII amacrine, we have also studied the membrane properties and inputs to rod bipolar cells, including the role of the glutamate transporter EAAT5 in regulating the release of glutamate from these cells (Veruki et al., 2006). Current projects include developing compartmental models for the different types of bipolar cells in the rat retina and for A17 amacrine cells.
Acknowledgements
We have had help along the way, so a big thank you to all the graduate students, postdocs and the technical team at the University of Bergen that have contributed to the work described above and all the other projects in the lab.
We first met in the laboratory of Professor Heinz Wässle at the Max Planck Institute for Brain Research, in Frankfurt, Germany. Heinz’s lab had just published the remarkable Boos et al., paper (1993) with the first patch-clamp recordings from a slice preparation of the mammalian retina, and we were both excited about the possibility to learn this technique. Eventually, we realized it was easier to join forces then compete with each other, and Meg ended up moving to Norway. The mystery of how the retina does what it does continues to propel us into the lab, even after all these years, to perform new experiments. AII amacrines, as well as their closest collaborators, still enchant us, and there are many more secrets to be revealed, so if you are looking for us, you will find us in the lab.
Key publications
- Liu JH, Singh JB, Veruki ML & Hartveit E (2021). Morphological properties of the axon initial segment-like process of AII amacrine cells in the rat retina. Journal of Comparative Neurology doi: 10.1002/cne.25210.
- Beltrán-Matas P, Hartveit E & Veruki ML (2021). Different glutamate sources and endogenous co-agonists activate extrasynaptic NMDA receptors on amacrine cells of the rod pathway microcircuit. European Journal of Neuroscience doi: 10.1111/ejn.15325
- Hartveit E, Veruki ML & Zandt B-J (2019). Capacitance measurement of dendritic exocytosis in an electrically coupled inhibitory retinal interneuron: an experimental and computational study. Physiological Reports e14186. doi: 10.14814/phy2.14186
- Veruki ML, Zhou Y, Castilho Á, Morgans CW & Hartveit E (2019). Extrasynaptic NMDA receptors on rod pathway amacrine cells: molecular composition, activation, and signaling. Journal of Neuroscience 39:627-650.
- Zandt B-J, Veruki ML & Hartveit E (2018). Electrotonic signal processing in AII amacrine cells: compartmental models and passive membrane properties for a gap junction-coupled retinal neuron. Brain Structure and Function 223: 3383-3410.
- Hartveit E, Zandt B-J, Madsen E, Castilho Á, Mørkve SH & Veruki ML (2018). AMPA receptors at ribbon synapses in the mammalian retina: kinetic models and molecular identity. Brain Structure and Function 223: 769-804.
- Zandt B-J, Liu JH, Veruki ML & Hartveit E (2017). AII amacrine cells: quantitative reconstruction and morphometric analysis of electrophysiologically identified cells in live rat retinal slices imaged with multi-photon excitation microscopy. Brain Structure and Function 222: 151-182.
- Zhou Y, Tencerová B, Hartveit E & Veruki ML (2016). Functional NMDA receptors are expressed by both AII and A17 amacrine cells in the rod pathway of the mammalian retina. Journal of Neurophysiology 115: 389-403
- Castilho Á, Madsen E, Ambrósio AF, Veruki ML & Hartveit E (2015b). Diabetic hyperglycemia reduces Ca2+ permeability of extrasynaptic AMPA receptors in AII amacrine cells. Journal of Neurophysiology 114: 1545-53.
- Castilho Á, Ambrósio AF, Hartveit E & Veruki ML (2015a). Disruption of a neural microcircuit in the rod pathway of the mammalian retina by diabetes mellitus. Journal of Neuroscience 35: 5422-5433.
- Hartveit E & Veruki ML (2012). Electrical synapses between AII amacrine cells in the retina: Function and modulation. Brain Research 1487: 160-172.
- Hartveit E & Veruki ML (2010) Accurate measurement of junctional conductance between electrically coupled cells with whole-cell voltage-clamp under conditions of high series resistance. Journal of Neuroscience Methods 187: 13-25.
- Veruki ML & Hartveit E (2009) Meclofenamic acid blocks electrical synapses of retinal AII amacrine and ON-cone bipolar cells. Journal of Neurophysiology 101: 2339-2347.
- Gill SB, Veruki ML & Hartveit E (2006). Functional properties of spontaneous IPSCs and glycine receptors in rod amacrine (AII) cells in the rat retina. Journal of Physiology 575: 739-759.
- Veruki ML, Mørkve SH & Hartveit E (2006). Activation of a presynaptic glutamate transporter regulates synaptic transmission through electrical signalling. Nature Neuroscience 9: 1388-1396.
- Veruki ML, Mørkve SH & Hartveit E (2003). Functional properties of spontaneous EPSCs and non-NMDA receptors in rod (AII) amacrine cells of the rat retina. Journal of Physiology 549: 759-774.
- Veruki ML & Hartveit E (2002b). Electrical synapses mediate signal transmission in the rod pathway of the mammalian retina. Journal of Neuroscience 22: 10558-10566.
- Veruki ML & Hartveit E (2002a). AII (rod) amacrine cells form a network of electrically coupled interneurons in the mammalian retina. Neuron 33: 935-946.
- Hartveit E & Veruki ML (1997). AII amacrine cells express functional NMDA receptors. NeuroReport 8: 1219-1223.
- Veruki ML & Wässle H (1996). Immunohistochemical localization of dopamine D1 receptors in rat retina. European Journal of Neuroscience 8: 2286-2297.
- Boos R, Schneider H & Wässle H (1993). Voltage- and transmitter-gated currents of all-amacrine cells in a slice preparation of the rat retina. Journal of Neuroscience 13: 2874-2888


Professors Meg Veruki and Espen Hartveit
Retinal Microcircuits Research Group
Department of Biomedicine
University of Bergen
Jonas Lies vei 91
N-5009 Bergen
Norway