vision-research.eu - The Gateway to European Vision Research
![]()

![]()
You are here: vision-research.eu » Vision Research » Visionary of the Quarter » Rigmor Baraas (Q01-2013)
Cone photoreceptors and visual sensitivity to colour, form and motion
 |
Advances in in-vivo retinal imaging techniques have made it possible to image individual photoreceptor cells in living humans. Adaptive optics imaging systems first measure, and then compensate for optical aberrations of the eye, permitting unprecedented detail of the living retina to be digitally captured. With the Kongsberg Adaptive Optics Ophthalmoscope, we combine high-resolution retinal imaging with experimental and clinical psychophysics and genetics to study normal and abnormal structural changes in the eye's photoreceptor layer and how this relates to perception of colour, spatial pattern and motion.
Our interest in cone photoreceptors and colour vision has developed from work on colour vision deficiencies and surface colour judgements. The incidence of inherited defects in colour vision is about 8-10% in the male population and about 0.5% in the female population (for recent results from screening of children in Norway please see Baraas, 2008). In the clinic and laboratory, the tasks of screening, grading, and diagnosis of the red-green deficiencies is straightforward, but these tests give a limited description of colour-vision function. We wanted to find out if colour-deficient observers are disadvantaged or not when it comes to perception of surface colour when viewing scenes imaged from the real world under different natural daylights. The objective was to test surface-colour perception with spectra drawn from natural scenes, but with no spatial cues (Baraas, et al. 2004), and to test this performance with natural scenes themselves (Baraas, et al. 2006, 2010). The results showed that in general red-green colour-deficient subjects were least disadvantaged with spectra drawn from natural scenes, even when these spectra were presented as part of geometric displays. Indeed, with these natural scenes, subjects with the most common colour deficiency (anomalous trichromacy) were able to discriminate natural surface colours under different lights as well as subjects with normal colour vision (Figure 1). Thus, clinical colour-vision tests may underestimate a subject's competence in judging surface colours in the natural world.
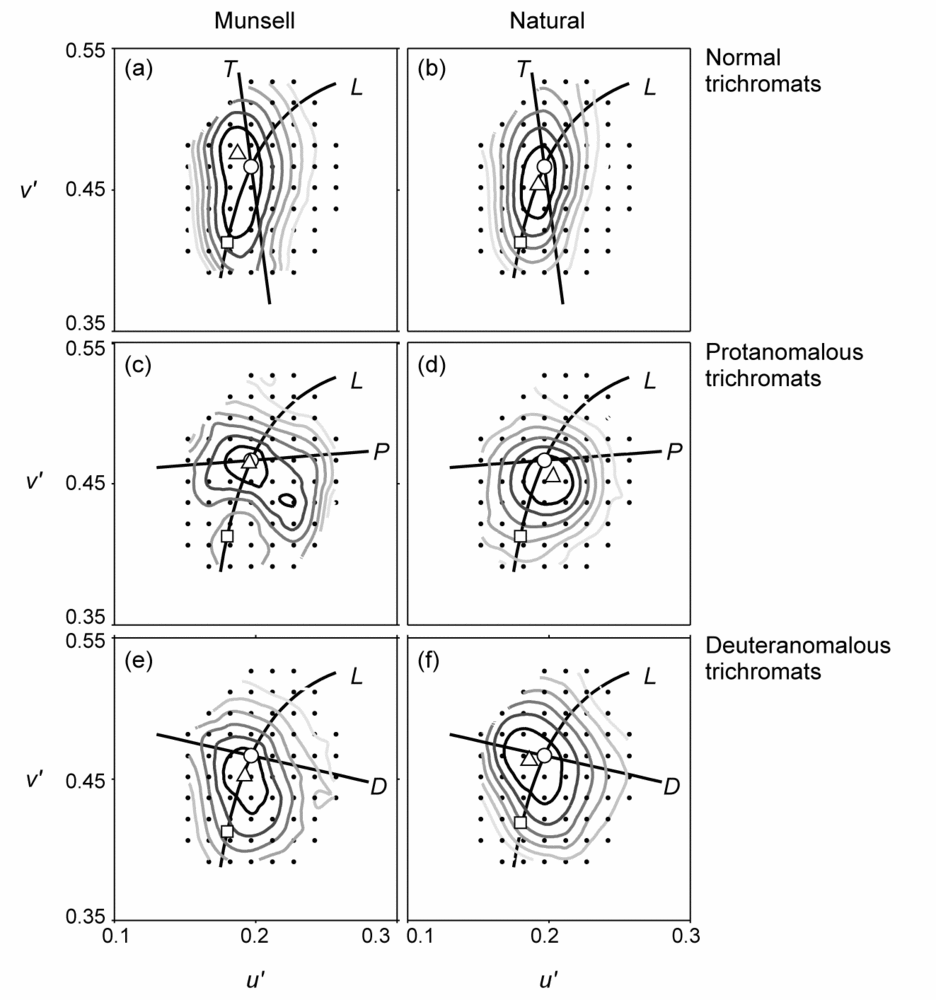 |
Over the last couple of years we have, in collaboration with Joseph Carroll at the Medical College of Wisconsin and Maureen Neitz at the University of Washington employed adaptive optics (AO) imaging to investigate the effects of certain cone photoreceptor opsin mutations on cone density and mosaic regularity. While it has been a common assumption that congenital colour deficiencies are stable throughout life, we have shown that inherited long-, medium- and short-wavelength colour-vision deficiencies can be non-stationary. Two patients with the Cys203Arg mutation on the OPN1MW gene were imaged to determine how the presence of this mutation affects the cones (Carroll, et al. 2009). Their colour vision phenotype was protanomalous and both had gene arrays with three OPN1MW genes and no OPN1LW genes. The Cys203Arg mutation was determined to be in the 2nd gene in one individual and the 3rd gene in the other. These first images of the Cys203Arg cone mosaic revealed a dramatic disruption, with about 40% reduction in cone density, providing direct evidence that expression of the Cys203Arg mutant pigment leads to degeneration of the associated cone photoreceptor. The reduction in cone density was accompanied by increased cone spacing with normal cone diameters. This disruption of the cone mosaic is consistent with degeneration of the cone cells that expressed the mutant pigment gene.
We have also discovered two novel mutations on the OPN1SW gene: R283Q and T190I. By the means of adaptive optics retinal imaging and retinal densitometry the R283Q mutation was shown to be progressive in nature, causing an S-cone dystrophy in the eldest person with the mutation (Baraas, et al. 2007). In a recent study we have investigated colour vision of subjects with the T190I mutation (Baraas, et al. 2012). Their performance was significantly different from that of normal trichromats along the tritan vector for both light-adapted and dark-adapted conditions and for both high and low levels of retinal illumination. Subjects with the T190I mutation behave as if they have S-cones that have reduced sensitivity at low light levels when compared with normal trichromats. This is the first mutation in extra-cellular loop II of the S-cone opsin gene that is associated with tritan colour-vision deficiency; the other five known mutations, including the R283Q mutation, are in the trans-membrane domains. An interesting aspect of the research is that the mutations that are non-stationary appear to follow what happens in analogous mutations in rhodopsin.
The Kongsberg Adaptive Optics Ophthalmoscope also allows us to study other types of cone disorders. Together with colleagues in Copenhagen, London and Milwaukee we examined the retinal structure in individuals with Oligocone trichromacy (OT) by the means of adaptive optics retinal imaging and optical coherence tomography. OT is an unusual cone dysfunction syndrome characterized by reduced visual acuity, mild photophobia, reduced amplitude of the cone electro-retinogram with normal rod responses, normal fundus appearance, and normal or near-normal colour vision. We showed that the foveal visual deficits in some individuals are caused by a reduced number of healthy cones (Michaelides, et al. 2011).
The advances in in-vivo imaging offer some exciting new opportunities to tie genetics and psychophysical results together with actual structural variation in the retina. We can use this to gain more knowledge about the organization of the photoreceptor mosaic in observers with normal and deficient colour or spatial vision. We can begin to understand the variation in cone density across different age groups and its impact on visual sensitivity to colour, form and motion. We can also examine correlates between structural changes in the eye's photoreceptor layer and visual perceptual changes in people with eyes that are affected by inherited and age-related retinal conditions.
 |
The retinal cone structure is instrumental in not only colour vision, but also spatial vision and motion processing. The large between-individual variability in cone density observed in the parafovea (Figure 2: Dees, et al. 2011), may have consequences with regards to normal and abnormal senescent changes in parafoveal visual function. Understanding variations in the cone photoreceptor mosaic and its implications on visual function will allow us to form a more coherent picture of the role played by the cone structure. This increased understanding could lead to the design of new and better vision tests which in turn could lead to earlier detection and intervention of retinal changes.
Selected References
- Baraas R. C., Hagen L. A., Dees E. W.,, Neitz M. (2012)
Substitution of isoleucine for threonine at position 190 of S-opsin causes S-cone-function abnormalities.
Vision Research 73, 1-9. - Dees E. W., Dubra, A., and Baraas R. C. (2011)
Variability in parafoveal cone mosaic in normal trichromatic individuals.
Biomedical Optics Express 2 (5), 1351-1358. - Michaelides M, Rha J, Dees EW, Baraas RC, Wagner-Schuman ML, Mollon JD, Dubis AM, Andersen MKG, Rosenberg T, Larsen M, Moore AT og Carroll J. (2011)
Integrity of the Cone Photoreceptor Mosaic in Oligocone Trichromacy.
Investigative Ophthalmology and Visual Science 52:4757-4764. - Baraas R. C., Foster D. H., Amano K., Nascimento S. M. C. (2010)
Color constancy of red-green dichromats and anomalous trichromats.
Investigative Ophthalmology and Visual Science. 51(4):2286-2293. - Carroll J, Baraas RC, Wagner-Schuman M, Rha J, Siebe CA, Sloan C, Tait DM, Thompson S, Morgan JI, Neitz J, Williams DR, Foster DH, Neitz M. (2009)
Cone photoreceptor mosaic disruption associated with Cys203Arg mutation in the M-cone opsin.
Proceedings of the National Academy of Sciences of the United States of America 106(49):20948-20953. - Baraas R. C. (2008)
Poorer color discrimination by females when tested with pseudoisochromatic plates containing vanishing designs on neutral backgrounds.
Visual Neuroscience. 25(3):501-505. - Baraas RC, Carroll J, Gunther KL, Chung M, Williams DR, Foster DH, Neitz M. (2007)
Adaptive optics retinal imaging reveals S-cone dystrophy in tritan color-vision deficiency.
Journal of the Optical Society of America 24(5):1438-1447. - Baraas R. C., Foster D. H., Amano K., Nascimento S. M. C. (2006)
Anomalous trichromats' judgments of surface color in natural scenes under different daylights.
Visual Neuroscience. 23(3-4):629-635. - Baraas R. C., Foster D. H., Amano K., and Nascimento S. M. C. (2004)
Protanopic observers show nearly normal color constancy with natural reflectance spectra.
Visual Neuroscience. 21(3), 347-351.
Professor Rigmor C Baraas
Director of Postgraduate Studies in Optometry and Visual Science
Head of Colour Vision and Retinal Imaging Research Group
Buskerud University College
Faculty of Health Sciences
Department of Optometry and Visual Science
Frogsvei 41
3611 Kongsberg, Norway
Phone: +47 32 86 97 87
E-mail: rigmor.baraas[at]hibu.no
Website: cvri.hibu.no