vision-research.eu - The Gateway to European Vision Research
![]()

![]()
You are here: vision-research.eu » Vision Research » Visionary of the Quarter » Thomas Langmann (Q01-2014)
Innate immunity and genetics in retinal degeneration
 |
Preface
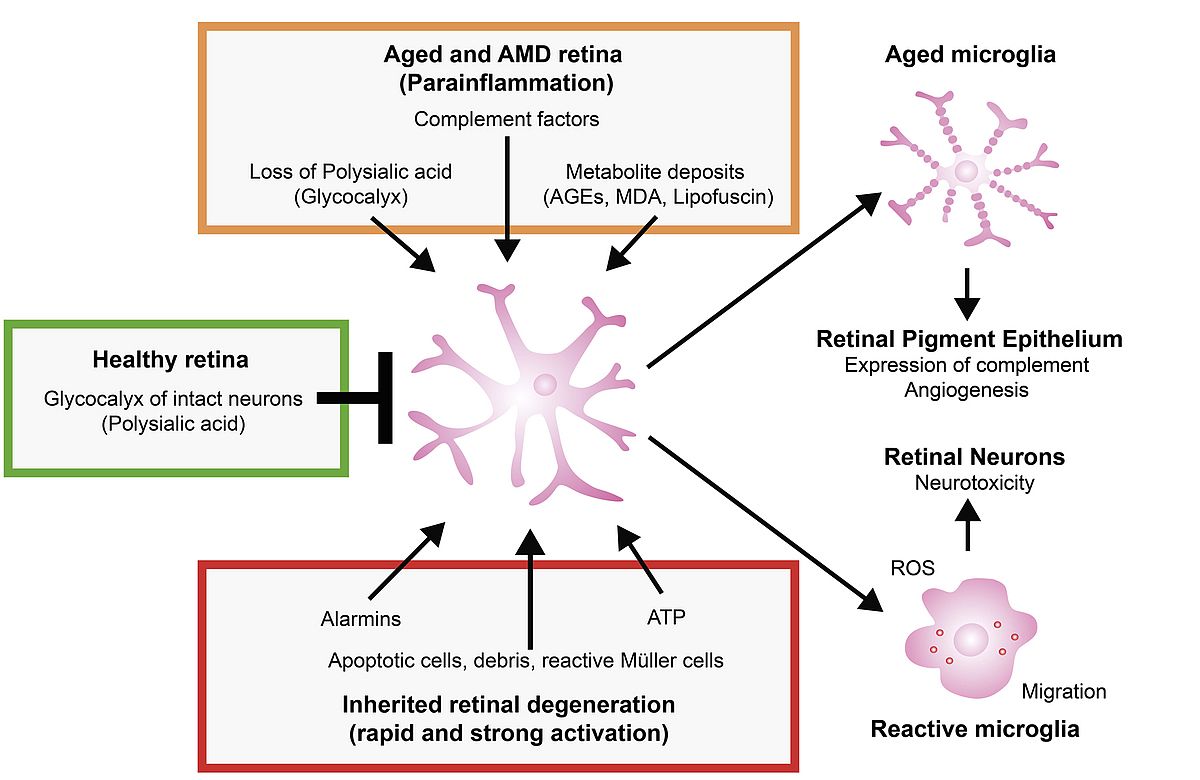
Activation of the innate immune system is an important hallmark of various retinal diseases including monogenic retinal dystrophies and age-related macular degeneration. The role of the complement factor system in AMD is now well documented by genetic associations of common and rare variants in complement genes and regulatory factors with AMD. However, the lack of good (dry) AMD animal models has hampered the systematic analysis of cellular immunity in the degenerating retina. In contrast, the reactivity of resident retinal microglial cells has been intensively characterized in several genetic and experimental mouse models for retinal degeneration. Based on data originating from our group and other laboratories over the last ten years, we postulate that i) mechanisms of immune cell reactivity are overlapping between rare inherited retinal degenerations and AMD, ii) resident glial cells, complement factors and RPE form a vicious circle of chronic inflammation in aged retinas, and iii) local therapeutic targeting of immune responses may be a strategy to support the survival of retinal neurons. Figure 1 summarizes the main features of our current concept.
 |
Microglial reactivity in retinal diseases
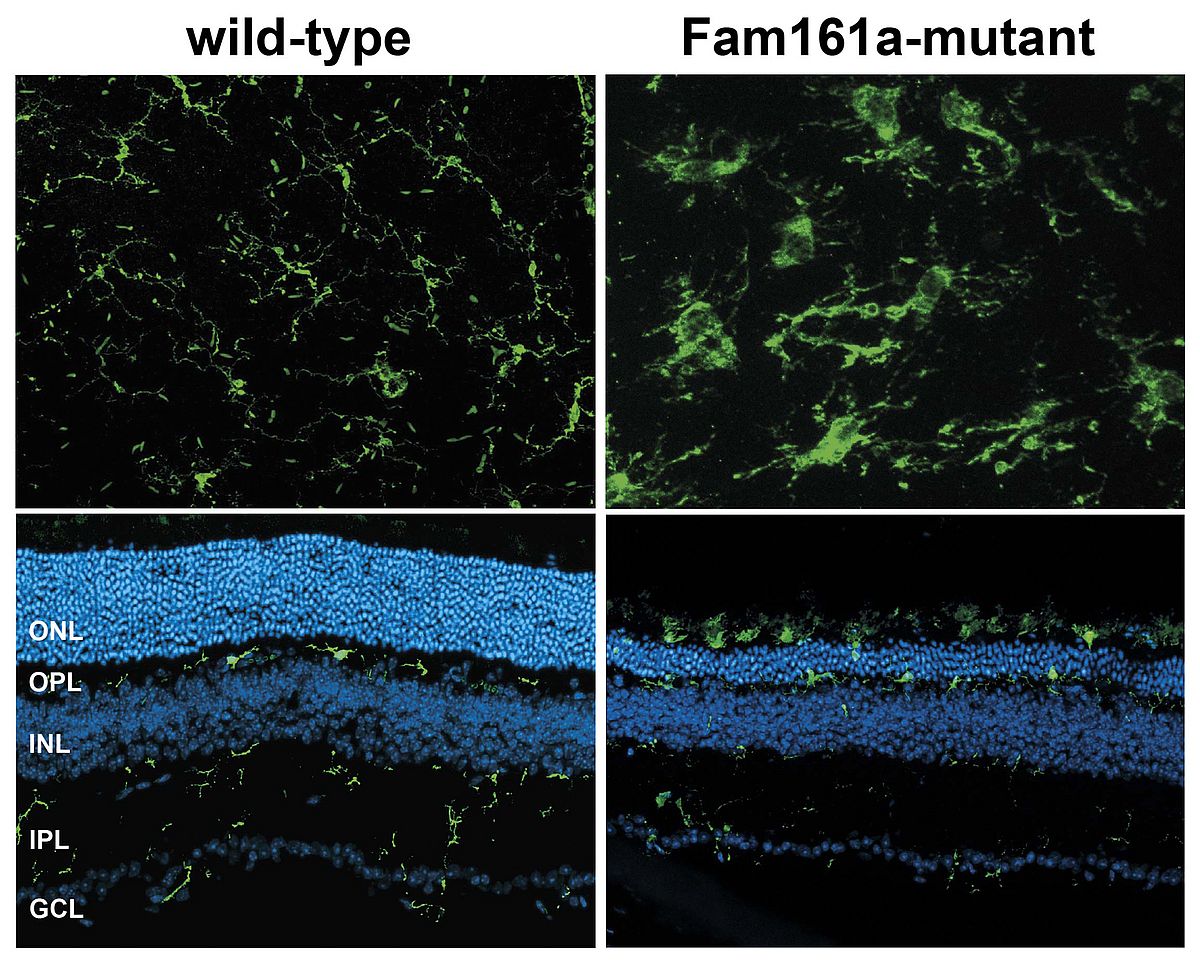
The focus of our laboratory is to understand why microglial cells are highly reactive to retinal damage and autonomous cell death. We have demonstrated that microglial cells are alerted before the onset of cell death in gene targeted mouse strains for retinoschisis, retinitis pigmentosa, and ceroid lipofuscinosis as well as in experimental models including light stress and laser-damage. In each of these models, microglial cells normally located in both plexiform layers of the retina, change their morphology from a highly ramified to an ameboid state and rapidly migrate to the site of damage (Figure 2).
 |
We have used microglia/macrophage GFP-reporter mice (MacGreen) to visualize and isolate microglial cells at different states of reactivity. Functional assays with cells isolated at the peak of disease progression clearly showed that they can actively produce neurotoxic reactive oxygen species, pro-inflammatory cytokines and inflammatory lipids. These detrimental consequences can be partially dampened by immuno-modulatory compounds including omega-3-fatty acids, curcumin, and luteolin. These microglia-modulating compounds then also have beneficial effects on retinal survival and attenuate retinal degeneration. Microglial cells can also produce endogenous self-regulatory factors such as ’activated microglia whey acidic protein’ (AMWAP), but mostly these local anti-inflammatory responses are overwhelmed by a latent parainflammation. We have also started to define molecular markers for reactive retinal microglia and could identify a strong expression of the mitochondrial translocator protein (18 kDa) (TSPO) in mouse and human retinas. Interestingly, pharmacological ligands such as XBD173 are available for this protein and first attempts using cell cultures and retinal explants showed a potent anti-inflammatory effect, encouraging a potential application in mouse models in vivo.
In very recent work with our colleague Harald Neumann (Bonn), we could show that a potential key component that triggers retinal microglial reactivity is a changed composition of the glycocalix on aged or damaged neurons. As final sugar cap of healthy neuronal glycoproteins, polysialic acid oligomers can efficiently bind to inhibitory microglial Siclec-receptors. Conversely, a loss of polysialic acid on retinal neurons can elicit a complement-receptor 3-mediated opsonization and pro-inflammatory activation of alerted microglial cells. We have therefore started to use polysialic acid treatment in the laser-model of choroidal neovascularization and first unpublished results are very promising. Polysialic acid-treated mouse eyes showed significantly reduced numbers of reactive microglial cells and displayed less vessel leakage and inflammation. Further work is now necessary to optimize the treatment protocol and to elucidate the molecular mechanisms responsible for the beneficial effects.
Retinal transcription factors and disease genetics
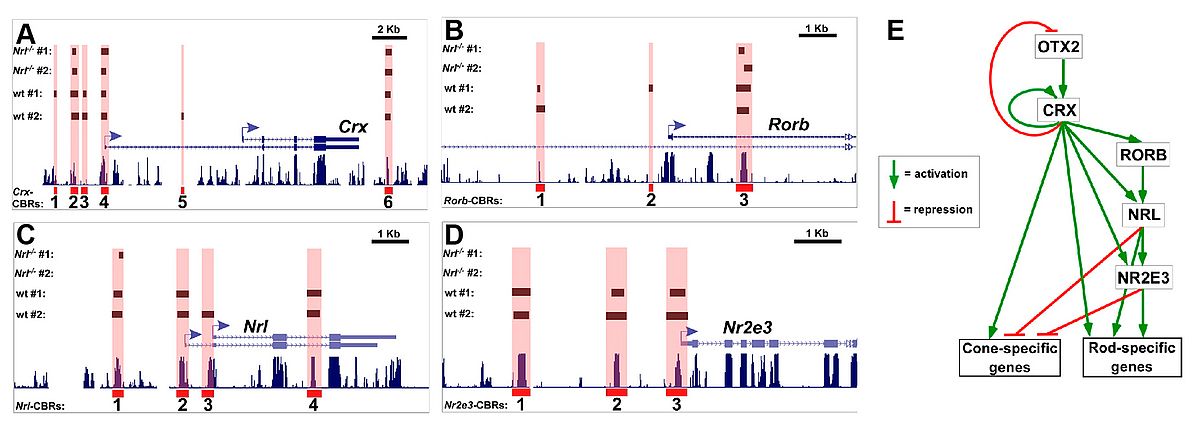
Another line of our research projects is related to retina-specific gene expression and identification of novel disease genes. Some years ago, we started to characterize transcriptional elements relevant for retinischisin transcription in the retina. Our data showed that the transcription factor ’cone-rod-homeobox (Crx) is crucial for retinoschisin expression in several species. Derived from this single locus analysis, we decided to go for a genome wide binding study using Crx ChIP-seq and were thereby the first to describe a full cistrome of a retinal transcription factor. Intriguingly, more than 5.000 genomic regions were bound by Crx in the mouse retina, including many orthologs of nearly all known human retinal disease genes. In a strategy termed ’cis-regulatory mapping’ we were able to prioritize and confirm the human FAM161A gene as causative gene for the retinitis pigmentosa 28 locus. We have also made our Crx ChIP-seq data set public and used it together with reporter electroporation into retinas to identify and characterize Samd7 as a novel transcriptional repressor in the mouse retina. Furthermore, Crx can be regarded as a retinal master transcription factor as it regulates many other transcription factors and thereby orchestrates a complex transcription factor regulatory network (Figure 3). Future projects in this area include the identification of other retinal transcription factor cistromes and their correlation with genetic variants related to AMD and rare inherited retinal dystrophies.
 |
Key publications from our laboratory
- Mirza M, Volz C, Karlstetter M, Langiu M, Somogyi A, Ruonala MO, Tamm ER,Jägle H, Langmann T. Progressive Retinal Degeneration and Glial Activation in the CLN6 (nclf) Mouse Model of Neuronal Ceroid Lipofuscinosis: A Beneficial Effect of DHA and Curcumin Supplementation. PLoS One. 2013 Oct 4;8(10):e75963
- Hlawatsch J, Karlstetter M, Aslanidis A, Lückoff A, Walczak Y, Plank M, Böck J, Langmann T. Sterile alpha motif containing 7 (samd7) is a novel crx-regulated transcriptional repressor in the retina. PLoS One. 2013;8(4):e60633.
- Ebert S, Walczak Y, Reme C, Langmann T. Microglial activation and transcriptomic changes in the blue light-exposed mouse retina. Adv Exp Med Biol. 2012;723:619-32.
- Karlstetter M, Lippe E, Walczak Y, Moehle C, Aslanidis A, Mirza M, Langmann T. Curcumin is a potent modulator of microglial gene expression and migration. J Neuroinflammation. 2011 Sep 29;8:125.
- Karlstetter M, Walczak Y, Weigelt K, Ebert S, Van den Brulle J, Schwer H, Fuchshofer R, Langmann T. The novel activated microglia/macrophage WAP domain protein, AMWAP, acts as a counter-regulator of proinflammatory response. J Immunol. 2010 Sep 15;185(6):3379-90.
- Langmann T, Di Gioia SA, Rau I, Stöhr H, Maksimovic NS, Corbo JC, Renner AB, Zrenner E, Kumaramanickavel G, Karlstetter M, Arsenijevic Y, Weber BH, Gal A, Rivolta C. Nonsense mutations in FAM161A cause RP28-associated recessive retinitis pigmentosa. Am J Hum Genet. 2010 Sep 10;87(3):376-81.
- Corbo JC, Lawrence KA, Karlstetter M, Myers CA, Abdelaziz M, Dirkes W, Weigelt K, Seifert M, Benes V, Fritsche LG, Weber BH, Langmann T. CRX ChIP-seq reveals the cis-regulatory architecture of mouse photoreceptors. Genome Res. 2010 Nov;20(11):1512-25.
- Dirscherl K, Karlstetter M, Ebert S, Kraus D, Hlawatsch J, Walczak Y, Moehle C, Fuchshofer R, Langmann T. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. J Neuroinflammation. 2010 Jan 14;7:3. doi: 10.1186/1742-2094-7-3.
- Ebert S, Weigelt K, Walczak Y, Drobnik W, Mauerer R, Hume DA, Weber BH, Langmann T. Docosahexaenoic acid attenuates microglial activation and delays early retinal degeneration. J Neurochem. 2009 Sep;110(6):1863-75.
- Langmann T, Lai CC, Weigelt K, Tam BM, Warneke-Wittstock R, Moritz OL, Weber BH. CRX controls retinal expression of the X-linked juvenile retinoschisis (RS1) gene. Nucleic Acids Res. 2008 Nov;36(20):6523-34.
- Ebert S, Schoeberl T, Walczak Y, Stoecker K, Stempfl T, Moehle C, Weber BH, Langmann T. Chondroitin sulfate disaccharide stimulates microglia to adopt a novel regulatory phenotype. J Leukoc Biol. 2008 Sep;84(3):736-40.
- Weigelt K, Ernst W, Walczak Y, Ebert S, Loenhardt T, Klug M, Rehli M, Weber BH, Langmann T. Dap12 expression in activated microglia from retinoschisin-deficient retina and its PU.1-dependent promoter regulation. J Leukoc Biol. 2007 Dec;82(6):1564-74.
Patent application:
Polysialic acid and use for treatment of neurodegenerative and neuroinflammatory diseases. Inventors: Neumann H, Kopatz J, Sharaz A, Karlstetter M, Langmann T.
Professor Thomas Langmann
Professor and Head
Experimental Immunology of the Eye
Email:
thomas.langmann[at]uk-koeln.de
Website:
augenklinik.uk-koeln.de/immunologie-des-auges
Contact
Department of Ophthalmology
University of Cologne
Building 35 (Anatomy)
Joseph-Stelzmann-Str. 9
50931 Cologne
Germany
Phone: +49 (0)221 478-7324
Fax: +49 (0)221 478-84591